For at det skal være en samsvar mellom informasjonen om polynukleotidet og det til polypeptidet, er det en kode: den genetiske koden.
De generelle egenskapene til den genetiske koden kan listes opp som følger:
Den genetiske koden består av trillinger, og er uten indre tegnsetting (Crick & Brenner,).
Det "ble dechiffrert ved bruk av" åpne celle -oversettelsessystemer "(Nirenberg & Matthaei, 1961; Nirenberg & Leder, 1964; Korana, 1964).
Det er sterkt degenerert (synonymer).
Organiseringen av kodetabellen er ikke tilfeldig.
Trillinger "tull".
Den genetiske koden er "standard", men ikke "universell".
Når vi ser på tabellen med den genetiske koden, må det huskes at den refererer til oversettelsen av "RNAm til polypeptid, som de involverte nukleotidbaser er A, U, G, C. Biosyntesen til en polypeptidkjede er oversettelsen av nukleotidsekvensen i sekvens aminosyre.

Hver basistriplett av RNAm, kalt kodon, har den første basen i venstre kolonne, den andre i den øverste raden, den tredje i den høyre kolonnen. La oss ta for eksempel tryptofan (dvs. Try), og vi ser at det tilsvarende kodonet vil være, i rekkefølge, UGG. Faktisk inkluderer den første basen, U, hele eskeraden øverst; i dette identifiserer G boksen til høyre og den fjerde linjen i selve boksen, der vi finner skrevet Try. På samme måte kan vi for å syntetisere tetrapeptidet Leucine-Alanine-Arginine-Serína (symbolene Leu-Ala-Arg-Ser) finne kodonene UUA-AUC-AGA-UCA i koden.

På dette tidspunktet bør det imidlertid bemerkes at alle aminosyrene i tetrapeptidet vårt er kodet (i motsetning til tryptofan) av mer enn ett kodon. Det er ikke tilfeldig at vi i det nettopp rapporterte eksemplet har valgt de angitte kodonene.Vi kunne ha kodet det samme tripeptidet med en annen RNAm-sekvens, for eksempel CUC-GCC-CGG-UCC.
I utgangspunktet fikk det faktum at en enkelt aminosyre tilsvarte mer enn en trilling en mening med tilfeldighet, også uttrykt i valget av begrepet degenerasjon av koden, som ble brukt for å definere fenomenet synonymi. På den annen side tyder noen data på at tilgjengeligheten av synonymer som kan refereres til ulik stabilitet i den genetiske informasjonen ikke er tilfeldig. Dette ser ut til å bli bekreftet også ved å finne en annen verdi av A + T / G + C -forholdet i de forskjellige evolusjonstrinnene. For eksempel, i prokaryoter, der behovet for variabilitet ikke er tilfredsstilt av reglene for mendelisme og neo-mendelisme, har A + T / G + C-forholdet en tendens til å øke. Den påfølgende lavere stabiliteten, i møte med mutasjoner, gir større muligheter for variabilitet tilfeldig fra genmutasjon.
I eukaryoter, spesielt i flercellede celler, der det er nødvendig at cellene i den enkelte organismen alle har samme arvelige arv, har A + T / G + C -forholdet i DNA en tendens til å redusere, noe som reduserer muligheten for somatiske genmutasjoner .
Eksistensen av synonyme kodoner i den genetiske koden reiser problemet, som allerede er nevnt, med mangfoldet eller ikke av antikodoner i RNAt.
Det er sikkert at det er minst ett RNAt for hver aminosyre, men det er ikke like sikkert om et enkelt RNAt kan binde seg til et enkelt kodon, eller kan gjenkjenne synonymer likegyldig (spesielt når disse er forskjellige bare for den tredje basen).
Vi kan konkludere med at det i gjennomsnitt er tre synonyme kodoner for hver aminosyre, mens antikodoner er minst en, og ikke mer enn tre.
Når vi husker at gener er ment som enkeltstrekninger av veldig lange polynukleotidsekvenser av DNA, er det klart at begynnelsen og slutten av det enkelte genet nødvendigvis må være inneholdt i minnet.
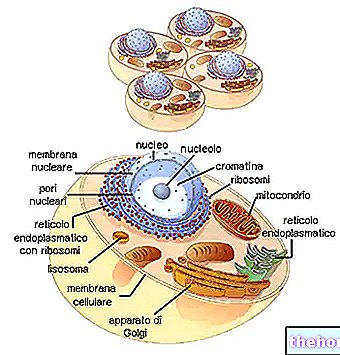
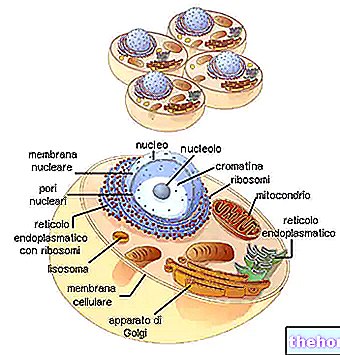
BIOSYNTESE AV PROTEINER
I forskjellige deler av DNA er det åpningen av dobbeltkjeden og syntesen av de forskjellige typene RNA.
Under lastingstrinnet binder RNAt seg til aminosyrene (tidligere aktivert av ATP og av det spesifikke enzymet). Det biosyntetiske "maskineriet" klarer ikke å "korrigere" feil ladede tRNA.
RNAr deler seg deretter inn i de to underenhetene, og ved binding til de ribosomale proteiner gir det opphav til samling av ribosomene.
RNAmet, som passerer gjennom cytoplasma, binder seg til ribosomene og danner polysomet.Hvert ribosom, som strømmer på budbringeren, er gradvis vert for RNAt som er komplementær til de relative kodonene, tar aminosyrene og binder dem til polypeptidkjeden i formasjon.
Den relativt stabile RNAt kommer inn i sirkulasjonen igjen. Ribosomene brukes også igjen og frigjør det allerede monterte polypeptidet.
Budbringeren, mindre stabil fordi det hele er monokatenært, spaltes (av ribonukleasen) inn i de konstituerende ribonukleotidene.
Syklusen fortsetter dermed og syntetiserer etter hverandre polypeptidene på messenger -RNA -ene levert av transkripsjonen.









-cos-cause-e-disturbi-associati.jpg)