NOTER OM EMBRYOLOGI
Embryologi studerer sekvensen av utviklingsformer fra zygoten til organismen utstyrt med alle dens organer og systemer.
I denne forbindelse er det verdt å huske skillet mellom utvikling (rekkefølge av strukturelle og organisatoriske faser med økende kompleksitet) og vekst, først og fremst ment i kvantitativ forstand.
I virveldyr metazoans vi er vitne til, stigende i den evolusjonære serien opp til mennesket (gjennom cyclostomes, fisk, amfibier, reptiler, fugler og pattedyr), utseendet av voksne former for økende kompleksitet, som komplikasjonen av stadier av embryonal utvikling.
I begynnelsen er zygoten, alltid utstyrt med reservemateriale, delt (ved påfølgende mitose) i 2, deretter 4, deretter 8 osv. Celler kalt blastomerer, uten vekst, til den når det normale kjerne / cytoplasmiske forholdet til arten .
Denne første segmenteringen kan følge forskjellige mønstre, avhengig av mengden og fordelingen av deutoplasma.
I begynnelsen er deutoplasma knapp ("oligolecytic egg"), så segmenteringen er total og gir opphav til svært forskjellige blastomerer. Etter hvert som kompleksiteten til embryoet øker, tar det mer tid og materiale før utviklingen tillater det å begynne. selvstendig liv. Dette krever en økning i deutoplasma ("telolecytiske egg"), som har en tendens til å ordnes i en del av zygoten. Dette forårsaker en økende "anisotropi", som er knyttet til modifikasjoner av segmenteringen, styrt av to generelle prinsipper:
- Hertwigs lov sier at ved mitose har den achromatiske spindelen (hvis ekvator bestemmer delingsplanet til dattercellene) en tendens til å være arrangert i retning av cytoplasmaets største lengde;
- Balfours lov sier at segmenteringshastigheten er omvendt proporsjonal med mengden deutoplasma.
Vi ser da at segmenteringen allerede i syklostomer og i fisk er ulik, med en raskt segmentert dyrestang (som vil gi embryoets øvre strukturer) og en eggeplomme som vil inneholde det meste av reservematerialet. Denne tendensen er enda større . anisotrop hos amfibier (der det er nødvendig å forberede organene som er ansvarlige for luftånding), der eggeplommestangen, mens den sakte segmenteres, forblir relativt inaktiv og ender med å bli dekket av celler som stammer fra den raskt segmenterte dyrepolen. rekkefølgen av de viktigste embryonale stadiene inkluderer: zygote, blastomerer, morula (klynge av blastomerer som ligner et bjørnebær), blastula (morula med regressive indre celler), gastrula (blastula der cellene på den ene siden har blitt invaginert), der det primitive hulrommet av organismen, med et eksternt cellelag (ektoderm, som nervesystemet først og fremst vil stamme fra) og en int erno (entoderm), mellom hvilket et tredje lag (mesoderm) deretter vil flytte seg. Fra disse lagene eller "embryonale ark" vil deretter, i en ordnet sekvens, utlede alt vev, organer og systemer.
I enda mer utviklede arter er økningen i deutoplasma (eller "kalv") slik at den ikke engang kan segmenteres. Vi ser dermed at segmentering hos fugler påvirker bare en tynn overfladisk skive, noe som fører til en "discoblastula" og en serie med fenomener som garanterer dannelsen av embryoet på en annen måte enn det som er nevnt ovenfor.
En ytterligere økning i deutoplasma ville sannsynligvis ikke vært mer effektiv, så hos pattedyr oppnås utvikling og vekst opp til evnen til selvstendig liv med et annet system.Vi bemerker faktisk hos pattedyr at deutoplasma bare brukes i de aller første stadiene av utvikling; da etablerer embryoet metabolske forhold til moderorganismen (med utseendet til morkaken) og bruker ikke lenger deutoplasma, hvis overskudd elimineres. På dette tidspunktet går eggene tilbake til å være oligolecitic og segmenteringen kan gå tilbake til å være total (og derfor i de tidlige stadiene ligner den på "amphioxus"), men etter morulaen fortsetter embryogenesen i henhold til det mest utviklede fuglemønsteret, med en "blastocyst" etterfulgt av implantasjon på livmorveggen, slik at metabolismen av embryoet sikres av moderorganismen (via morkaken) i stedet for av deutoplasma.
EMBRYO -DIFFERENSJON
Når segmenteringen av zygoten har brakt kjerne / cytoplasmatisk forhold til artens norm, må veksten også begynne, parallelt med utviklingen.Derfor begynner metabolismen, med utseendet av nukleoli og proteinsyntese. Proteinsyntesen som dermed initieres skyldes genene som er ansvarlige for de tidlige stadiene av embryonal utvikling. Disse genene er deprimert av stoffene som er tilstede i de forskjellige blastomerene på dyret og kalvpolen. På sin side kan produktene av disse tidlige genene deprimere operonene til genene som har ansvaret for de senere stadiene. Produktene fra denne andre serien av gener vil kunne virke både i betydningen å konstruere nye embryonale strukturer og i den forstand å undertrykke de forrige operonene og derepressere de følgende, i en ordnet sekvens som fører til konstruksjonen av den nye organismen , takket være den genetiske informasjonen som er akkumulert. fra genomet gjennom årtusener til stadig mer utviklede arter.
Haeckels berømte uttrykk "" ontogeny summerer fylogeni "uttrykker faktisk det faktum at de høyere artene gjentar, i stadiene av embryonal utvikling, den rekkefølgen som allerede er funnet i de evolusjonært tidligere artene.
De tidlige stadiene av embryoet har en tendens til å være like hos virveldyr, spesielt opp til gjellens utseende.
Hos arter som bytter til luftånding, blir gjellene deretter absorbert og gjenbrukt (for eksempel for dannelse av endokrine kjertler), men den genetiske informasjonen knyttet til dannelsen av gjellene er også bevart hos mennesker. Dette er åpenbart et eksempel på embryonale strukturelle gener som er tilstede i genomet til alle virveldyr og må forbli undertrykt etter å ha fungert i sitt ontogenetiske øyeblikk.
Tolkningen av embryogenese i betydningen regulering av genhandling gjør det mulig å forene de komplekse tradisjonelle erfaringene med eksperimentell embryologi.
TVILLINGER
Zygoten og de første blastomerene, inntil proteinsyntesen begynner, er totipotente, det vil si i stand til å gi liv til en hel organisme. Til dette er eksperimentene til Spemann knyttet, som skaffet to embryoer fra kvelning av en amfibisk zygote. Et lignende fenomen vises på grunnlag av fenomenet identiske tvillinger hos mennesker, som nettopp av denne grunn kalles monozygotisk (MZ). Spemanns eksperimentelle tvillinger var halve størrelsen på normal, mens de hos mennesker er helt normale. Dette forklares fordi i to amfibier måtte de to embryoene dele den eneste eggeplommen som allerede er mottatt, mens embryoene hos mannen kan motta alt som er nødvendig for deres utvikling og vekst gjennom morkaken.
Det skal huskes at hos "mennesker har to tredjedeler av tilfellene av tvillinger" en annen opprinnelse: de stammer fra en og annen modning av to follikler samtidig, med frigjøring av to eggløsninger som, når de befruktes, gir to zygoter; faktisk, i i dette tilfellet snakker vi om dizygotiske tvillinger (DZ).
Siden MZ -tvillingene, delt med mitose fra den eneste zygoten, har samme genom, må forskjellene mellom dem være av miljømessig opprinnelse.I stedet er genomet til to DZ -tvillinger bare like mye som for to brødre. Tvillingmetode, mye brukt i menneskelig genetikk og også innen sport.
I "mannen, der visse etiske årsaker ville forby eksperimentering, kan det fastslås hvor mye en karakter er regulert av arvelige faktorer: Faktisk er strengt arvelige karakterer (for eksempel blodgrupper) alltid samsvarende bare hos MZ tvillinger; at de samsvar med et trekk i MZ er nær det til DZ, det er utledet at miljøfaktorer råder over arvelige faktorer ved å bestemme den fenotypiske egenskapen.

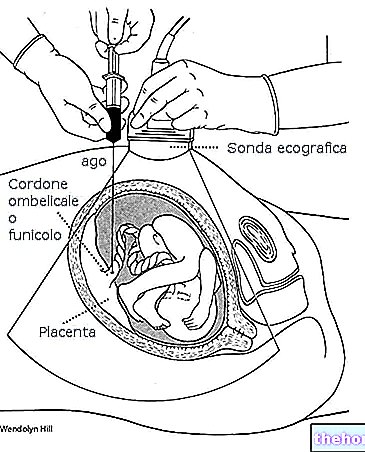
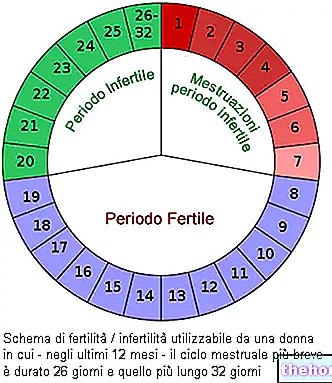

-cos-come-funziona-a-cosa-serve.jpg)



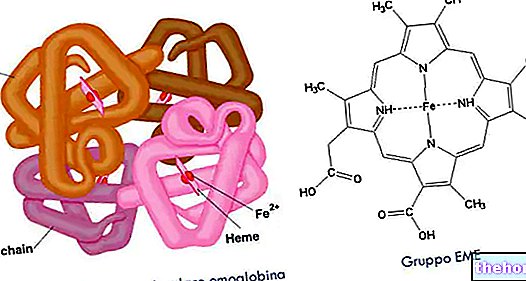










-cause-sintomi-e-terapia.jpg)



