Definisjon
Enzymer er proteiner produsert i plante- og dyreceller, som fungerer som katalysatorer som akselererer biologiske reaksjoner uten å bli modifisert.
Enzymer virker ved å kombinere med et bestemt stoff for å omdanne det til et annet stoff; klassiske eksempler er gitt av fordøyelsesenzymer som er tilstede i spytt, mage, bukspyttkjertel og tynntarm, som utfører en viktig funksjon i fordøyelsen og hjelper til med å bryte ned mat til grunnleggende bestanddeler, som deretter kan absorberes og brukes av kroppen, behandlet av andre enzymer eller skilles ut som avfall.
Hvert enzym har en bestemt rolle: Det som bryter ned fett, virker for eksempel ikke på proteiner eller karbohydrater. Enzymer er avgjørende for organismens velvære. Mangel, selv på et enkelt enzym, kan forårsake alvorlige forstyrrelser. Et velkjent eksempel er fenylketonuri (PKU), en sykdom preget av manglende evne til å metabolisere en essensiell aminosyre, fenylalanin, hvis opphopning kan forårsake fysiske misdannelser og psykiske sykdommer.
Biokjemisk studie
Enzymer er spesielle proteiner som har karakteristikken for å være biologiske katalysatorer, det vil si at de har evnen til å bryte ned aktiveringsenergien (Eatt) til en reaksjon, og modifiserer dens vei slik at en kinetisk sakte prosess er raskere.
Enzymer øker kinetikken til termodynamisk mulige reaksjoner, og i motsetning til katalysatorer er de mer eller mindre spesifikke: de har derfor substratspesifisitet.
Enzymet er ikke involvert i reaksjonens støkiometri: for at dette skal skje må det siste katalytiske stedet være identisk med det som starter.
I den katalytiske virkningen er det nesten alltid en langsom fase som bestemmer hastigheten på prosessen.
Når vi snakker om enzymer er det ikke riktig å snakke om likevektsreaksjoner, vi snakker i stedet om stabil (tilstand der en viss metabolitt dannes og forbrukes kontinuerlig, og holder konsentrasjonen nesten konstant over tid). Produktet av en reaksjon katalysert av ett enzym er vanligvis selv en reaktant for en påfølgende reaksjon, katalysert av et annet enzym, og så videre.
Prosesser katalysert av enzymer består vanligvis av sekvenser av reaksjoner.
En generisk reaksjon katalysert av et enzym (E) kan oppsummeres som følger:

E er enzymet
S er underlaget;
ES representerer adduktet mellom enzym og substrat;
P er produktet;
K er reaksjonens hastighetskonstant.
Et generisk enzym (E) kombineres med substratet (S) for å danne adduktet (ES) med en hastighetskonstant K1; det kan dissosiere tilbake til E + S, med en hastighetskonstant K2, eller, (hvis "lever" lenge nok ) kan fortsette til form P med en hastighetskonstant K3.
Produktet (P) kan igjen rekombinere med enzymet og reformere adduktet med hastighetskonstant K4.
Når enzymet og substratet blandes, er det en brøkdel av tiden der møtet mellom de to artene ennå ikke har skjedd: det vil si at det er et ekstremt kort tidsintervall (som avhenger av reaksjonen) der enzym og substrat har ennå ikke oppfylt; etter denne perioden kommer enzym og substrat i kontakt i økende mengder og ES -adduktet dannes. Deretter virker enzymet på substratet og produktet frigjøres. Det kan da sies at c "er et innledende tidsintervall der konsentrasjonen av ES -adduktet ikke kan defineres; etter denne perioden antas det at en stabil tilstand er etablert, det vil si at hastigheten på prosessene som fører til å få adduktet er lik hastigheten på prosessene som fører til ødeleggelse av adduktet.
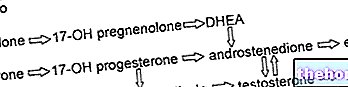
Michaelis-Menten-konstanten (KM) er en likevektskonstant (referert til den første likevekten beskrevet ovenfor); det kan sies, med en god tilnærming (fordi K3 også bør vurderes), at KM er representert ved forholdet mellom de kinetiske konstantene K2 og K1 (referert til ødeleggelse og dannelse av adduktet ES i den første likevekten beskrevet ovenfor) .
Gjennom Michaelis-Menten-konstanten har vi en "indikasjon på affiniteten mellom enzym og substrat: hvis KM er liten c" er en "høy affinitet mellom enzym og substrat, så er ES-adduktet stabilt.
Enzymer er underlagt regulering (eller modulering).
Tidligere var det hovedsakelig snakk om negativ modulering, det vil si inhibering av et enzyms katalytiske evner, men det kan også være en positiv modulasjon, det vil si at det er arter som kan forbedre et enzyms katalytiske evner.
Det er 4 typer hemninger (hentet fra tilnærminger gjort på en modell for å matche eksperimentelle data med matematiske ligninger):
- konkurransehemming
- ikke-konkurransedyktig hemming
- inkompetitiv inhibering
- konkurransehemmende
Vi snakker om konkurransehemming når et molekyl (hemmer) er i stand til å konkurrere med underlaget. For strukturell likhet kan inhibitoren reagere i stedet for substratet, derav terminologien "konkurrerende inhibering". Sannsynligheten for at enzymet binder seg til inhibitoren eller substratet avhenger av konsentrasjonen av begge og deres affinitet med enzymet; reaksjonshastigheten avhenger derfor av disse faktorene.
For å oppnå samme reaksjonshastighet som uten tilstedeværelse av inhibitoren, er det nødvendig å ha en høyere substratkonsentrasjon.
Det er eksperimentelt vist at i nærvær av en hemmer øker Michaelis-Menten-konstanten.
Når det gjelder i stedet "den ikke-konkurrerende inhiberingen, samspillet mellom molekylet som skal fungere som en modulator (positiv eller negativ hemmer) og" enzymet, finner sted på et sted som er forskjellig fra det der interaksjonen er forekommer mellom enzym og substrat; Vi snakker derfor om allosterisk modulering (fra gresk allosteros → annet nettsted).
Hvis inhibitoren binder seg til enzymet, kan det indusere en endring i enzymets struktur og kan følgelig redusere effektiviteten som substratet binder seg til enzymet med.
I denne typen prosesser forblir Michaelis-Menten-konstanten konstant siden denne verdien avhenger av likevekten mellom enzymet og substratet, og selv i nærvær av en hemmer endres ikke disse likevektene.
Fenomenet inkompetitiv inhibering er sjeldent; en typisk inkompetitiv hemmer er et stoff som reversibelt binder seg til ES -adduktet som gir opphav til ESI:

Inhibering fra substratoverskudd kan noen ganger være inkompetitiv, da dette skjer når et andre substratmolekyl binder seg til ES -komplekset, noe som gir opphav til ESS -komplekset.
En konkurrerende inhibitor kan derimot bare binde seg til substratenzymadduktet som i det forrige tilfellet: bindingen av substratet til det frie enzymet induserer en konformasjonsmodifikasjon som gjør stedet tilgjengelig for inhibitoren.
Michaelis Menten -konstanten synker når hemmerkonsentrasjonen øker: tilsynelatende øker derfor enzymets affinitet for substratet.
Serin protease
De er en familie av enzymer som chymotrypsin og trypsin tilhører.
Chymotrypsin er et proteolytisk og hydrolytisk enzym som skjærer til høyre for hydrofobe og aromatiske aminosyrer.
Produktet av genet som koder for kymotrypsin er ikke aktivt (det aktiveres med en kommando); den inaktive formen av kymotrypsin er representert av en polypeptidkjede på 245 aminosyrer. Chymotrypsin har en kuleform på grunn av fem disulfidbroer og andre mindre interaksjoner (elektrostatisk, Van der Waals -krefter, hydrogenbindinger, etc.).
Chymotrypsin produseres av chymosecellene i bukspyttkjertelen der det er inneholdt i spesielle membraner og blir utvist gjennom bukspyttkjertelen i tarmen på tidspunktet for fordøyelse av mat: chymotrypsin er faktisk et fordøyelsesenzym. Proteinene og næringsstoffene vi inntar gjennom dietten utsettes for fordøyelse for å reduseres til mindre kjeder og absorberes og omdannes til energi (f.eks. Amylaser og proteaser bryter ned næringsstoffer til glukose og aminosyrer som når cellene, gjennom blodårene de når portalvenen og blir derfra transportert til leveren hvor de gjennomgår videre behandling).
Enzymer produseres i en ikke-aktiv form og aktiveres bare når de når "stedet der de må operere"; når handlingen er fullført, deaktiveres de. Et enzym, når det først er deaktivert, kan ikke reaktiveres: for å ha en "ytterligere katalytisk virkning, må det erstattes av" et annet enzymmolekyl. Hvis chimitrypsin ble produsert i aktiv form allerede i bukspyttkjertelen, ville det angripe sistnevnte: pankreatitt er patologier på grunn av fordøyelsesenzymer som allerede er aktivert i bukspyttkjertelen (og ikke på de nødvendige stedene); noen av dem hvis de ikke blir behandlet i tide, føre til døden.
I kymotrypsin og i alle serinproteaser skyldes den katalytiske virkningen eksistensen av alkoholanionen (-CH2O-) i sidekjeden til en serin.
Serinproteaser tar dette navnet nettopp fordi deres katalytiske virkning skyldes en serin.
Når alt enzymet har utført sin handling, før det kan operere på underlaget igjen, må det gjenopprettes med vann; "frigjøringen" av serin ved vannet er den tregeste fasen av prosessen, og det er denne fasen som bestemmer hastigheten på katalyse.
Den katalytiske virkningen skjer i to faser:
- dannelse av anionen med katalytiske egenskaper (anionalkoholat) og påfølgende nukleofilt angrep på karbonylkarbonet (C = O) med spaltning av peptidbindingen og dannelse av esteren;
- vannangrep med restaurering av katalysatoren (i stand til å utøve sin katalytiske virkning igjen).
De forskjellige enzymene som tilhører serinproteasefamilien kan bestå av forskjellige aminosyrer, men for dem alle er det katalytiske stedet representert av det alkoholiske anionen i sidekjeden til en serin.
En underfamilie av serinproteaser er enzymene som er involvert i koagulering (som består i transformasjon av protein, fra deres inaktive form til en "annen form som er aktiv). Disse enzymene sikrer at koagulering er så effektiv som mulig og er begrenset i plass og tid (koagulering må skje raskt og må bare forekomme i nærheten av det skadede området). Enzymene som er involvert i koagulering aktiveres i en kaskade (fra aktivering av et enkelt enzym, oppnås milliarder av enzymer: hvert aktivert enzym , i sin tur aktiverer mange andre enzymer).
Trombose er en patologi på grunn av funksjonssvikt i koaguleringsenzymer: det er forårsaket av aktivering, uten behov (fordi det ikke er noen skade), av enzymene som brukes i koagulering.
Det er modulerende (regulatoriske) enzymer og hemmende enzymer for andre enzymer: interaksjon med sistnevnte, de regulerer eller hemmer aktiviteten; selv produktet av et enzym kan være en hemmer for enzymet. Det er også enzymer som virker jo mer, jo større er substratet.
Lysozym
Luigi Pasteur oppdaget ved nysing på en petriskål at i slimet er det et enzym som er i stand til å drepe bakterier: lysozym; fra gresk: liso = hvilken størrelse; zimo = enzym.
Lysozym er i stand til å bryte ned celleveggen til bakterier. Bakterier og encellede organismer generelt trenger mekanisk resistente strukturer som begrenser formen; inne i bakteriene er det et veldig høyt osmotisk trykk, slik at de tiltrekker seg vann. Plasmamembranen ville eksplodere hvis det ikke var noen cellevegg som motsetter inntrengning av vann og begrenser volumet av bakterien.
Celleveggen består av en polysakkaridkjede der molekyler av N-acetyl-glukosamin (NAG) og molekyler av N-acetyl-muraminsyre (NAM) veksler; bindingen mellom NAG og NAM brytes ved hydrolyse.Karboksylgruppen til NAM, i celleveggen, er engasjert i en peptidbinding med en aminosyre.
Mellom de forskjellige kjedene dannes broer som består av pseudo-peptidbindinger: forgreningen skyldes lysinmolekylet; strukturen som helhet er veldig forgrenet, og dette gir den en høy stabilitet.
Lysozym er et antibiotikum (dreper bakterier): det virker ved å lage en sprekk i bakterieveggen; når denne strukturen (som er mekanisk motstandsdyktig) bryter, trekker bakterien vann til den sprenger. Lysozym klarer å bryte β-1,4 glukosidbinding mellom NAM og NAG.
Det katalytiske stedet for lysozym er representert av et spor som går langs enzymet der polysakkaridkjeden settes inn: seks glukosidiske ringer i kjeden plasseres i sporet.
I posisjon tre av sporet c "er en choke: i denne posisjonen kan bare én NAG plasseres, fordi NAM, som har høyere dimensjoner, ikke kan komme inn. Det faktiske katalytiske stedet er mellom posisjoner fire og fem: siden det er en NAG i posisjon tre, vil kuttet finne sted mellom en NAM og en NAG (og ikke omvendt); kuttet er derfor spesifikt.
Den optimale pH -verdien for lysozym å fungere er fem. På enzymets katalytiske sted, dvs. mellom posisjonene fire og fem, er det sidekjedene til en asparaginsyre og en glutaminsyre.
Grad av homologi: måler slektskap (dvs. likhet) mellom proteinstrukturer.
Det er et sterkt forhold mellom lysozym og laktosesyntase.
Laktosesyntetase syntetiserer laktose (som er det viktigste melkesukkeret): laktose er et galaktosylglukosid der c "er en β-1,4 glukosidbinding mellom galaktose og glukose.
Derfor katalyserer laktosesyntetase den motsatte reaksjonen til den som katalyseres av lysozym (som i stedet deler β-1,4 glukosidbindingen)
Laktosesyntetase er en dimer, det vil si at den består av to proteinkjeder, hvorav den ene har katalytiske egenskaper og er sammenlignbar med lysozym, og den andre er en regulatorisk underenhet.
Under graviditet syntetiseres glykoproteiner av cellene i brystkjertelen ved virkningen av galatosyl-tranferase (den har en "sekvenshomologi på 40% med lysozym): dette enzymet er i stand til å overføre en galaktosylgruppe fra en høyenergistruktur til en glykoproteinstruktur. Under graviditet induseres ekspresjonen av genet som koder for galaktosisyl-transferase (det er også uttrykk for andre gener som også gir andre produkter): det er en økning i brystets størrelse fordi det er aktivert brystkjertelen (tidligere inaktiv) som må produsere melk Under fødsel produseres α-laktalbumin som er et regulatorisk protein: det er i stand til å regulere katalytisk kapasitet til galaktosyltransferase (ved diskriminering av underlaget). Galaktosyltransferase modifisert av α-laktalalbumin er i stand til å overføre en galaktosyl til et glukosemolekyl: danner en β-1,4 glykosidbinding og gir laktose (laktosesyntetase).
Derfor forbereder galaktose -transferase brystkjertelen før levering og produserer melk etter levering.
For å produsere glykoproteiner, binder galaktosyltransferase seg til en galaktosyl og en NAG; under fødsel binder laktalalbumin seg til galaktosyltransferase, noe som gjør at sistnevnte gjenkjenner glukose og ikke lenger NAG for å gi laktose.