FORENING (ELLER "CONCATENATION" ELLER "LINKAGE")
Hittil har vi behandlet den mendelske dihybrid (eller polyhybrid) kryssingen forutsatt at de forskjellige allelparene faktisk finnes på forskjellige homologe kromosompar. Men antallet kromosomale par, selv om det er forskjellig fra art til art, varierer innenfor trange grenser (få arter når rundt hundre kromosomer), mens antall gener kan telles i titusenvis.
At tegnene som Mendel valgte for sine eksperimenter segregerte alle uavhengig (uten derfor å forvirre beregningene i fordelingen av fenotyper i F2 på polyhybridkorset) var en heldig sjanse. Hvis to par alleler ble funnet på tilstøtende loci, var loven som ville følge ville bli kalt loven.
Når man vet at veldig mange tegn har sitt lokus på et enkelt kromosom og at de er de kromosomale parene som skiller seg uavhengig av hverandre i meiose, kan det godt forstås hvor ofte det skjer at to par tegn, hvis de var assosiert på et kromosom hos foreldrene organisme, forblir assosiert. også i kjønnscellen og derfor i organismen som den vil bringe sitt eget genetiske materiale til.
Dermed ser vi at "assosiasjon representerer et" unntak, langt fra sjeldent, til uavhengigheten i Mendels tredje lov.
Bytte eller "krysse-over" og rekombinasjon
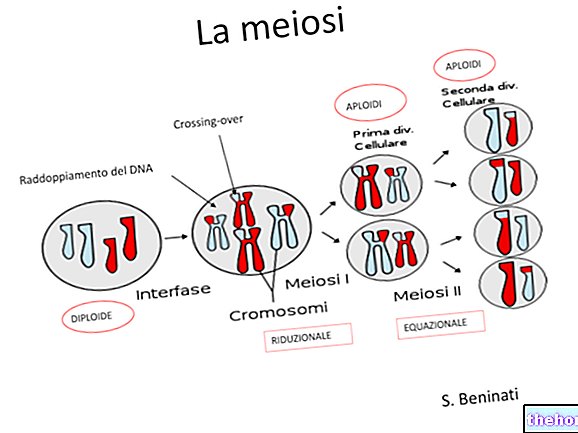
Når vi snakker om meiose, har vi indikert at det er to forskjellige øyeblikk for blanding av det genetiske materialet: det ene er segregeringen av kromosomer i kjønnsceller, og det er det som ble observert av Mendel.
Det andre øyeblikket, som faktisk går foran, er det der de fire kromatidene i hvert par homologe kromosomer utveksler identiske egenskaper. Etter denne utvekslingen vil to faktorer som var assosiert på det samme kromosomet i stedet være uavhengige i kjønnscellene. Sannsynligheten at en utveksling finner sted er proporsjonal, med en første tilnærming, til lengden på kromosomet, og i lengre kromosomer kan det være enda mer enn en utveksling.
Fenomenet kan oppdages cytologisk, ved å observere et tilstrekkelig antall meioser under mikroskopet.
Re-match rate er frekvensen hvor to tegn som var assosiert i foreldrenes generasjon, rekombinerer forskjellig i F2.
Hvis de to loci er absolutt sammenhengende, vil sannsynligheten for at en chiasme vil skille dem være praktisk talt null. Rekombinasjonshastigheten vil være: n ° rekombinanter. Hvis to loci er på to forskjellige kromosomer, vil rekombinasjonshastigheten være 0,5 (like sannsynlighet, for to tegn som ble slått sammen i P -generasjonen, for å tilfeldig finne seg sammen i F2). Rekombinasjonshastigheten kan derfor variere mellom 0,0 og 0,5. For små avstander på kromosomet er avstanden og rekombinasjonshastigheten direkte proporsjonal. For lengre distanser er det mulighet for at to utvekslinger finner sted mellom to loci. Det vil nå fremstå som klart at to faktorer atskilt med to børser igjen er forbundet. det er klart, på dette tidspunktet, at proporsjonaliteten mellom avstanden til lokalene og sannsynligheten for rekombinasjon er tapt.
Loci funnet assosiert på det samme kromosomet utgjør 'assosiasjonsgrupper'. Svært fjerne loci kan ha en slik sannsynlighet for separasjon ved utveksling at de oppfører seg som uavhengige, men hver av dem vil bli assosiert, med en lavere rekombinasjonshastighet, til mellomliggende loci.
Når rekombinasjonshastigheten mellom mange par gener i en assosiasjonsgruppe er kjent, kan konstruksjonen av 'genetiske kart' begynne. Husk at avstanden mellom to gener (a og b) uttrykkes av rekombinasjonshastigheten og at avstanden til a fra et tredje gen c kan være enten summen eller forskjellen med hensyn til avstanden fra b, er det mulig å rekonstruere et kart over de gjensidige avstandene, som vil være det genetiske kartet innenfor den gruppen av assosiasjoner, det vil si det kromosomet.
Vi må nå generelt vurdere noen begreper som begrenser den fenotypiske manifestasjonen av genotypiske tegn.
Først av alt vil vi snakke om begrepene penetrasjon og uttrykksevne, og deretter vil vi vie spesiell oppmerksomhet til fenomenene regulering av genhandling.
PENETRANCE
Penetranse av et gen representerer dets evne til å manifestere seg i fenotypen. Gjennomtrengelighet måles statistisk ved å telle frekvensen av fenotypene som viser denne karakteren av 100 genotyper som inneholder den. En egenskap med 0,7 penetrasjon er en egenskap som forekommer fenotypisk i 70% av dens genotypiske frekvens.
EKSPRESSIVITET
Ekspressivitet er en kvantitativ vurdering av graden av fenotypisk manifestasjon.
FORORDNING OM GENEHANDLING
Celler produserer alle enzymer og proteiner med samme hastighet og samtidig. Escherichia coli-celler kan for eksempel tilføres energi og karbonatomer fra laktosedisakkaridet ettersom de er i stand til å bryte dem ned til glukose og galaktose takket være beta-galaktosidaseenzymet. I et normalt E. coli som kan ha laktose, det er omtrent 3000 molekyler av beta-galaktosidase, lik 3% av proteinene i den cellen; i fravær av laktose vil det bare være ett molekyl beta-galaktosidase per bakteriecelle. Galaktosidase vil bli syntetisert fra nye mRNA -molekyler når den kan brukes. Mutantstammer av E. coli som er rike på enzymet er kjent selv når laktose er fraværende: disse mutantene er dårligere sammenlignet med normale celler ettersom de blir tvunget til unødvendig forbruk av energi og materialer for å produsere enzymet som vil forbli uten substrat. Stoffer som forårsaker en økning i mengden enzym, som tilfellet er med laktose, vil bli kalt induktorer, mens enzymer sies å være induserbare. Andre stoffer induserer, også disse på en bestemt måte, produksjonen av visse enzymer. Også i E. coli, for eksempel i stand til å bygge alle sine aminosyrer, med karbon og ammonium (NH3), blokkerer tilstedeværelsen i kulturmediet av en bestemt aminosyre (for eksempel histidin) produksjonen av alle enzymer assosiert med biosyntesen til selve aminosyren: det vil bli sagt om disse enzymene at de er repressible. I bakterieceller blir mRNA -molekylene revet kort tid etter dannelsen, og det er derfor å kontrollere produksjonen av mRNA betyr å kontrollere den enzymatiske syntesen samtidig tid.
OPERONEN
For å forklare hvordan bakteriecellen er i stand til å kontrollere sin egen produksjon av enzymer, formulerte Jacob og Monod hypotesen om operonet; operon dannes av flere gener som er funksjonelt beslektet og justert uten diskontinuitet langs en strekning av DNA. Operonet består av tre forskjellige typer gener: promotoren, der dannelsen av mRNA begynner; operatøren, hvor kontroll utøves; ett eller flere strukturelle gener, som koder for enzymer eller for andre proteiner. I beta-galaktosidasesystemet inkluderer operonet, i tillegg til det for beta-galaktosidase, også to andre gener strukturell koding for andre enzymer involvert i metabolismen av laktose. Disse genene er tilstøtende til hverandre og transkriberes etter hverandre langs samme DNA -helix til et enkelt mRNA -molekyl. De således produserte mRNA -molekylene er aktive i svært kort tid, hvoretter de blir ødelagt av spesifikke enzymer..
Operonens aktivitet styres igjen av et annet gen, regulatoren, som også kan være fjernt fra operonet: denne regulatoren koder for et protein, kalt en repressor, som ser ut til å binde seg til DNA ved operatørgenet. mellom promotoren og de strukturelle genene blokkerer faktisk produksjonen av mRNA.
Repressoren kontrolleres igjen, og kontrollen utføres ved hjelp av et "signal" stoff. Når det gjelder induserbare enzymer er dette stoffet "induktoren. Induktoren" binder seg til at repressormolekylet endrer formen slik at den ikke lenger kan tilpasse seg DNA: i dette tilfellet, siden det ikke er noen repressor mellom promotoren og strukturelle gener , kan repressoren danne mRNA -molekylene og fra disse proteinmolekylene. Med utmattelse av tilførsel av induktor igjen vil regulatoren gjenvinne kontrollen, noe som vil stoppe produksjonen av nytt mRNA, derfor av nye proteiner.I beta-galaktosidasesystemet er induktoren laktose eller et stoff som ligner veldig på dette. Derivat: de vil slutte seg til repressoren og inaktivere den for å tillate biosyntese av enzymer. Når det gjelder repressible enzymer, virker stoffet som fungerer som et "signal" som en corepressor: repressoren er bare aktiv hvis den kombineres med corepressoren. I histidinsystemet, som involverer et dusin forskjellige enzymer, er det denne aminosyren, kombinert med dens tRNA, corepressor. Histidine.
ALLOSTERISKE INTERAKSJONER
Allosteriske interaksjoner, som involverer inaktivering av et enzym ved å endre dets form, gir en annen måte å regulere den metabolske aktiviteten til en celle. Allosteriske interaksjoner gir mulighet for mer nøyaktig kontroll enn induktor -repressorsystemet til operonet, men oppnår ikke det nyttige resultatet ved å ekskludere biosyntesen til et gitt stoff fra det første trinnet - produksjonen av et mRNA.
KONTROLLSYSTEMER I EUCARIUS
Det er noen fakta som kan anta at et reguleringssystem som ligner operonen fungerer og er fremtredende blant planter og dyr.Kromosomene til disse organismene skiller seg sterkt fra E. coli og andre prokaryoter. Kontrollen av genene i disse celler er veldig forskjellige. Mekanismen for mitose er slik at hver celle i en gitt plante eller et dyr besitter all informasjon
genetikk tilstede i det befruktede egget. Derfor vil de fleste genene i enhver spesialisert celle forbli ineffektiv gjennom cellens levetid. DNA i disse cellene er alltid assosiert med proteiner. Så det er mulig at genrepresjon i eukaryoter krever nettopp denne sammenhengen mellom DNA og proteiner.



















-cos-cause-sintomi-e-cura.jpg)