Mendel, Gregor - bohemsk naturforsker (Heinzendorf, Schlesien, 1822 -Brno, Moravia, 1884). Etter å ha blitt en augustinsk munter, gikk han inn i klosteret i Brno i 1843; deretter fullførte han sine vitenskapelige studier ved Universitetet i Wien. Fra 1854 underviste han i fysikk og naturvitenskap i Brno. Mellom 1857 og 1868 viet han seg til lange praktiske eksperimenter på hybridisering av erter i klosterhagen. Etter nøye og tålmodig observasjon av resultatene, ble han ledet til med klarhet og matematisk nøyaktighet å angi viktige lover som går under navnet Mendels lover. Disse lovene var like gyldige for planteverdenen som for dyrene og utgjorde utgangspunktet for etableringen av en ny gren av biologiske vitenskaper: genetikk. I ni år, mens han analyserte resultatene av hundrevis og hundrevis av kunstige pollinasjoner, dyrket og undersøkte rundt 12 000 planter, registrerte Mendel tålmodig alle observasjonene, hvis resultater ble presentert i en kort memoar til Brno Natural History Society i 1865. På tiden ble publikasjonen ikke verdsatt i all sin betydning og vakte ikke "interessen den fortjente. Ignorert av lærde i mer enn tretti" år, ble lovene gjenoppdaget i 1900 samtidig og uavhengig av tre botanikere: H. de Vries i Holland , C. Currens i Tyskland, E. von Tschermak i Østerrike; men i mellomtiden hadde studiet av biologi gjort store fremskritt, tider hadde endret seg og oppdagelsen hadde umiddelbart stor innvirkning.
Den første loven, eller loven om dominans, kalles også mer riktig loven om ensartethet av hybrider. Mendel tok to erteplanter (som han kalte forfedre) begge av ren rase, den ene med gule frø, den andre grønne, og brukte pollen til den ene for å gjødsle den andre. Fra dette korset en første generasjon erter av hybridplanter, som ikke lenger er renavlet, avledet; alle plantene produserte gulfrøede erter, ingen viste grønnfrøkarakteren. Den gule skriften dominerte med andre ord det grønne; det vil si at gul var dominerende, grønn, maskert, recessiv. Det er også et spesielt tilfelle når det er ufullstendig dominans og den første generasjonen viser en mellomliggende karakter mellom det farlige og det morlige; men selv i dette tilfellet vil hybrider være lik hverandre. Mendel ga en strålende og genial forklaring på fenomenene; han antok at faktorer som var ansvarlige for utvikling av karakterer ble overført sammen med kjønnscellene; han trodde at i hver organisme eller en gitt karakter er regulert av to faktorer, en overført av moren og en av faren, og at disse to faktorene er de samme hos renrasede individer, forskjellige i hybrider, og at til slutt bare en faktor er inneholdt i kjønnsceller. Mendel angav de to faktorene til de antagonistiske tegnene med bokstaver i alfabetet, store bokstaver for dominerende, små bokstaver for recessive; og siden hver forelder har et par faktorer, indikerte han for eksempel med AA erten som bærer den dominerende gule karakteren, med aa den som bærer den grønne recessive karakteren. Hybriden, som mottar A fra den ene forelderen og fra den andre, vil være Aa.
Her kan det påpekes at ut fra et individs utseende er det ikke alltid mulig å vite om den tilhører en ren rase eller om den er en hybrid; i stedet er det nødvendig å undersøke dens oppførsel i interbreeding og crossbreeding. Faktisk er de renrasede gule erter og hybridene tilsynelatende identiske; Imidlertid er det kjent at deres genetiske sammensetning er forskjellig, den ene er AA og den andre Aa. Når du krysser mellom dem rase gule erter (AA), vil du alltid og bare ha gulfrøede erter, krysse gule eller halvgule, men hybridærter (Aa) med hverandre, og du vil også se planter med grønne frø dukke opp etterkommerne deres . De gule erter Aa, selv om de er identiske, er forskjellige genotypisk, det vil si i deres genetiske sammensetning. Andre viktige lover i Mendel er: loven om adskillelse eller disjunksjon av karakterer og loven om karakterers uavhengighet.
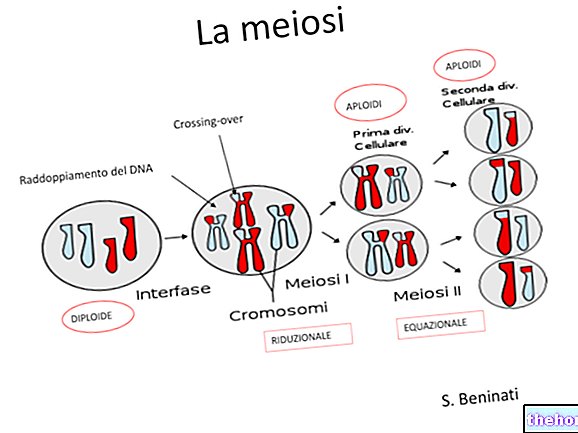
På Mendels tid var fenomenene mitose og meiose ennå ikke klare, men i dag vet vi at i meiosen mottar kjønnscellene bare ett kromosom av hvert par, og at disse kromosomene utelukkende med befruktning kommer tilbake for å parre seg tilfeldig.
Hvis vi tror (for midlertidig forenkling) at en bestemt faktor er lokalisert på et enkelt kromosompar, ser vi at faktorene i den eukaryote (diploide) organismen er parvis, og bare i kjønnscellene (haploide) er det en enkelt faktor. der de er tilstede i par, kan de enten være like eller forskjellige.
Når to like faktorer (enten dominerende eller recessive, GG eller gg) har slått seg sammen til zygoten, sies det resulterende individet å være homozygot for den karakteren, mens heterozygot er en der to forskjellige faktorer har slått seg sammen (Gg).
De alternative faktorene som bestemmer karakteren i individet kalles alleler. I vårt tilfelle er henholdsvis G og g den dominerende allelen og den recessive allelen for erternes fargekarakter.
Allelene for en bestemt karakter kan være enda mer enn to. Vi vil derfor snakke om dialleliske og polialleliske tegn, eller henholdsvis om genetisk dimorfisme og polymorfisme.
Etter konvensjon er generasjonene av det eksperimentelle korset angitt med symbolene P, F1 og F2, som henholdsvis betyr:
P = foreldrenes generasjon;
F1 = første filiale generasjon;
F2 = andre generasjon filial.
I det mendelske korset gir gult X grønt alle gule; to av de sistnevnte, krysset med hverandre, gir en grønn for hver tre gule. De gule og grønne i generasjon P er alle homozygote (som fastslått med et langt utvalg). de gir alltid like kjønnsceller, så deres avkom er like like, alle heterozygoter Siden gul er dominerende over grønn, er heterozygoter alle gule (F1).
Ved å krysse to av disse heterozygotene ser vi imidlertid at hver enkelt kan gi den ene eller den andre typen gameter med like stor sannsynlighet. Også foreningen av gameter i zygotene har samme sannsynlighet (unntatt i spesielle tilfeller), slik at i F2 dannes zygoter av de fire mulige typene med like sannsynlighet: GG = homozygot, gul; Gg = heterozygot, gul; gG = heterozygot, gul; gg = homozygot, grønn.
Gult og grønt er derfor i et forhold på 3: 1 i F2, ettersom gult forekommer uansett så lenge det er tilstede, mens grønt bare forekommer i fravær av gult.
For bedre å forstå fenomenet fra molekylærbiologisk synspunkt, er det tilstrekkelig å anta at et bestemt grunnstoff, grønt, ikke modifiseres av enzymet produsert av g -allelet, mens G -allelet produserer et enzym som omdanner det grønne pigment til gult pigment. Hvis G -allelen ikke er tilstede på noen av de to homologe kromosomene som bærer det genet, forblir ertene grønne.
Det faktum at gule erter kan preges av to forskjellige genetiske strukturer, den homozygote GG og heterozygoten Gg, gir oss muligheten til å definere fenotypen og genotypen.
Den ytre manifestasjonen av organismens genetiske egenskaper (det vi ser), mer eller mindre modifisert av miljøpåvirkninger, kalles fenotype. Settet med genetiske egenskaper alene, som kanskje eller ikke kan manifesteres i fenotypen, kalles genotype.
F2 gule erter har samme fenotype, men variabel genotype. Faktisk er de 2/3 heterozygoter (bærere av den recessive egenskapen) og 1/3 homozygoter.
I stedet, for eksempel, i grønne erter er genotypen og fenotypen gjensidig uforanderlige.
Som vi vil se, er utseendet til bare en av foreldrekarakterene i F1, og utseendet til begge tegnene i et 3: 1 -forhold i F2, fenomener av generell karakter som er gjenstand for Mendels henholdsvis 1. og 2. lov. Alt dette refererer til kryssingen mellom individer som er forskjellige for et enkelt par alleler, for en enkelt genetisk karakter.
Hvis det foretas en annen slik kryssing, gjentas det mendelske mønsteret; for eksempel ved å krysse erter med rynket frø og glatt frø, der den glatte allelen er dominerende, vil vi ha LL X 11 i P, alle LI (heterozygote, glatte) i F1 og tre glatte for hver rynket i F2 (25 % LL, 50% LI, 25% 11). Men hvis vi nå krysser homozygote dobler, det er varianter som er forskjellige med mer enn ett tegn (for eksempel GGLL, gult og glatt, med ggll, grønt og regosi), ser vi at i F1 vil alle være heterozygote med begge dominerende karakterer, fenotypet, men i F2 vil det være de fire mulige fenotypiske kombinasjonene i et numerisk forhold på 9: 3: 3: 1 som stammer fra de 16 mulige genotypene som tilsvarer de mulige kombinasjonene av fire typer gameter (tatt i par til to i zygotene).
Det er tydelig at to karakterer som var sammen i den første generasjonen uavhengig av hverandre skiller hverandre i den tredje. Hvert par homologe kromosomer segregerer uavhengig av det andre ved meiose, og dette er hva Mendels tredje lov fastslår.
La oss nå se som en helhet en formulering av Mendels tre lover:
1a: dominanslov. Gitt et par alleler, hvis avkommet til et kryss mellom de respektive homozygotene bare har en av foreldrekarakterene i fenotypen, kalles dette dominant og den andre recessiv.
2a: lov om segregering. Kryssing av F1 -hybrider gir tre dominanter for hver recessiv.Fenotypisk forhold er derfor 3: 1, mens det genotypiske forholdet er 1: 2: 1 (25% dominerende homozygoter, 50% heterozygoter, 25% recessive homozygoter).
Når man krysser individer som er forskjellige med mer enn ett par alleler, segreger hvert par i etterkommere, uavhengig av de andre, i henhold til 1. og 2. lov.
Disse tre lovene, selv om de ikke er riktig formulert som sådan av Mendel, er anerkjent som grunnlaget for eukaryotisk genetikk. Som det alltid er tilfelle i biologiens store prinsipper, betyr ikke den generelle karakteren til disse lovene at de ikke har noen unntak.
Det er faktisk så mange mulige unntak at det i dag er vanlig å dele genetikk inn i mendelsk og neomendelsk, inkludert i sistnevnte alle fenomener som ikke faller innenfor mendelsk lov.
Mens de første unntakene imidlertid tviler på gyldigheten av Mendels oppdagelser, var det senere mulig å demonstrere at lovene hans er generelle i omfang, men de underliggende fenomenene kombineres med en rekke andre fenomener som modulerer dem. Ellers uttrykket.
FORTSETT: Forutsi barnets blodgruppe "