Uten genetisk variasjon bør alle levende ting (ved arv) være det samme som det første. For å ha ulik vesener vil de eneste forklaringene være de som er knyttet til enkeltkreasjoner. Men vi vet at strukturen til DNA, som er grunnlaget for overføring av arvelige tegn, har en relativ og ikke en absolutt stabilitet. Selv om stabiliteten garanterer bevaring av den første informasjonen, bestemmer ustabiliteten modifikasjonene, eller bedre (for å bruke det spesifikke uttrykket) mutasjonene.
Mutasjoner kan deles inn i 3 store grupper:
- genmutasjoner;
- kromosomale mutasjoner;
- genomiske mutasjoner.
På dette tidspunktet er det godt å kort legge til to begreper: det ene er "reparasjon" og det andre er "gunstig" eller "ugunstig" mutasjon.
Reparasjonen, et engelsk begrep som betyr reparasjon, refererer til eksistensen i cellene av spesifikke enzymatiske prosesser som er beregnet på å verifisere bevaring av informasjon, eliminere eller korrigere strekningene av DNA som ikke samsvarer med originalen.
Begrepet "gunstig" eller "ugunstig" mutasjon uttrykker sammenligningen mellom effektiviteten, for organismen og for arten, av det opprinnelige genet ("vill", dvs. "vill", primitiv) og av mutantgenet.
Det skal bemerkes at fordeler og ulemper alltid refererer til visse miljøforhold; en ugunstig mutasjon i et bestemt miljø kan bli gunstig under forskjellige forhold.
Konsekvensene av dette prinsippet er enorme i populasjonsgenetikk, som det allerede kan demonstreres av de forskjellige overlevelseseffektivitetene til hemoglobinvarianter i normale eller malarielle miljøer. Men dette emnet bør utsettes til mer detaljerte artikler om generell, menneskelig eller medisinsk genetikk.
Et annet skille er nødvendig (spesielt for genmutasjoner) mellom somatiske mutasjoner og germline mutasjoner. Alle cellene i vår flercellede organisme stammer som kjent fra en enkelt zygote, men veldig tidlig differensierer de til en somatisk linje, som hele kroppen utvikler seg fra, og en kimlinje som er bestemt til å danne gonadene og, for terminal meiose, gameter Det er ganske tydelig at, bortsett fra interaksjoner mellom de to cellepopulasjonene, vil en mutasjon av den somatiske linjen manifestere seg i den enkelte organismen, men vil ikke påvirke etterkommerne, mens en mutasjon i kimlinjen bare vil forekomme hos etterkommerne.
Når det gjelder de forskjellige mutasjonstyper og de relative konsekvensene, er det verdt å nevne en klassifisering av årsakene til mutasjon. Disse årsakene kalles mutagener, som hovedsakelig brukes til å skille mellom fysiske og kjemiske mutagener. Ulike modifikasjoner av det fysiske miljøet kan føre til mutasjoner, men de viktigste fysiske mutagenene er stråling.Av denne grunn er radioaktive stoffer farlige, og det er fremfor alt de tyngre radioaktive partiklene som har en tendens til å forårsake mutasjoner på grunn av avføring, med de alvorligste konsekvensene.
Kjemiske mutagener kan fungere både ved å endre den ordnede strukturen til nukleinsyrer, og ved å introdusere stoffer som ligner normale nitrogenholdige baser i celler, som kan konkurrere med sistnevnte i syntesen av nukleinsyrer, og dermed forårsake mutasjoner ved substitusjon.
GENE MUTASJONER
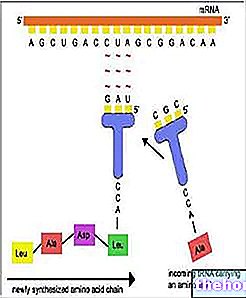
Genmutasjoner påvirker ett eller noen få gener, det vil si en begrenset DNA -strekning. Siden informasjon lagres i en sekvens av nukleotidpar, består den minste mutasjonsenheten (et muton) av et enkelt komplementært basepar.Uten å gå i detalj om de forskjellige mutasjonsmekanismene på gennivå, kan vi begrense oss til å nevne to: basesubstitusjon og gjenvalg (eller innsetting). Ved basesubstitusjonsmutasjoner erstattes ett eller flere DNA -nukleotider med andre. Hvis feilen ikke blir korrigert i tide, vil en sekvens følge på transkripsjonstidspunktet også endret i RNA. Hvis endringen av triplet er ikke begrenset til et synonym (se den genetiske koden), det vil resultere i substitusjon av en eller flere aminosyrer i polypeptidsekvensen. Substitusjonen av en aminosyre kan være mer eller mindre kritisk for bestemmelsen av proteinets struktur og dets funksjon.
Ved gjenvalg eller innsettingsmutasjoner fjernes ett eller flere nukleotider eller legges til DNA -sekvensen. Disse mutasjonene er generelt veldig alvorlige fordi (med mindre de er hele trillinger som legger til eller trekker fra enkelte aminosyrer) alle trillingene som følger i leserekkefølgen endres.
Genmutasjoner er de hyppigste og er opprinnelsen til de fleste variasjonene i arvelige egenskaper mellom individer.
KROMOSOMISKE MUTASJONER
Dette er mutasjoner som påvirker relativt lange fragmenter av et kromosom. Det er vanlig å klassifisere dem hovedsakelig i:
- kromosommutasjoner for gjenvalg;
- kromosommutasjoner ved duplisering;
- kromosommutasjoner ved translokasjon.
Mutasjoner på grunn av avføring oppstår på grunn av brudd og tap av et mer eller mindre langt fragment av kromosom. Spesielt ved meiose er denne typen mutasjoner ofte dødelig, på grunn av totalt tap av et visst antall gener, mer eller mindre uunnværlig.
Ved duplikasjonsmutasjoner, etter en pause, har kromatidstubbene en tendens til å smelte sammen.
I den påfølgende fjerningen av sentromerer bryter kromosomet, som har blitt dicentrisk, ofte i ulik deler: som vi kan se, er resultatet gjenvalg på den ene siden og duplisering på den andre.
Et kromosombrudd kan etterfølges av en reversering. Det totale genetiske materialet er uendret, men sekvensen til genene på kromosomet er endret.
Tilfellet med en translokasjon er lik, men det gjelder binding av et fragment av et kromosom til et ikke-homologt kromosom. Det ene kromosomet er amputert og det andre forlenget; den totale genetiske informasjonen til cellen er fortsatt uendret, men posisjonseffekten er enda mer markert. Det er lett å representere en posisjonseffekt ved å referere til begrepet regulering av genhandling: ved å endre posisjon på kromosomer, kan et gen lett forlate en operon og settes inn i en annen, noe som resulterer i en "endret aktivering eller undertrykkelse.
Imidlertid sies det at en translokasjon er balansert (eller balansert) når det er gjensidighet mellom translokasjon mellom to par kromosomer, og holder summen av geninformasjonen uendret. Den kryssformede figuren i meiotisk diakinesis tilsvarer generelt den balanserte translokasjonen.
GENOMISKE MUTASJONER
Når man husker at genomet er det individuelle genetiske arv, ordnet i kromosomer, kan det spesifiseres at vi snakker om genomiske mutasjoner når kromosomene har en annen fordeling enn normen for arten.
Genomiske mutasjoner kan hovedsakelig klassifiseres i mutasjoner for polyploidi, haploidi og aneuploidi.
Polyploidymutasjoner oppstår når reduksjon ikke resulterer i deling; de forekommer lettere i planter, der de faktisk brukes til å forbedre produksjonen.
Hvis celledeling mislykkes i meiose, kan det oppstå diploide gameter; hvis en slik kjønnscelle klarer å slå seg sammen med en normal kjønnscelle, vil zygoten som følge av denne befruktningen være triploide. En slik zygote kan noen ganger lykkes med å stamme fra en hel organisme, siden reduksjon og mitose ikke krever et jevnt antall kromosomer. På tidspunktet for meiose vil imidlertid regelmessig sammenkobling av homologe kromosomer være umulig.
Genomiske mutasjoner ved haploidi kan oppstå når en gamete i en normalt diploid art blir aktivert av en annen gamete blottet for kjernefysisk materiale, eller til og med i fravær av befruktning: et haploid individ vil oppstå.
Mens de tidligere typene genomiske mutasjoner alltid angår addisjon eller subtraksjon av hele tall n av kromosomer, gjelder mutasjoner for aneuploidi overskudd eller defekt av enkeltkromosomer (kromosomavvik).
Den euploide kromosomale sammensetningen av en art er definert som dens normale karyotype eller idiotype.